
引用本文: 林浩, 王朝樣, 张海宝, 巨家华, 俞永江. 微小RNA在胃癌干细胞干性调控及靶向治疗中的研究进展. 中国普外基础与临床杂志, 2023, 30(3): 358-363. doi: 10.7507/1007-9424.202209019 复制
胃癌是严重威胁人类健康的恶性肿瘤之一,全球每年新发病例约93万例,近70万人死亡,其发病率和病死率分别位居所有恶性肿瘤的第4位和第2位[1]。胃镜等临床诊断技术的进步提高了胃癌患者的早期检出率[2]。对于早期胃癌患者,手术切除有着良好的疗效。然而,对于晚期患者,由于存在局部浸润和远处转移,治疗选择仍然有限,手术切除效果不理想。进一步鉴定调控胃癌进展的关键分子可为晚期胃癌患者提供新的治疗靶点[3]。肿瘤干细胞(cancer stem cells,CSCs)是能够引起肿瘤生长并具有自我更新、多向分化能力的一类细胞亚群,可以分化成大量新的肿瘤细胞,与肿瘤的恶性生物学行为及不良预后有着重要的关系[4]。研究发现胃癌干细胞(gastric cancer stem cells,GCSCs)在胃癌疾病进展过程中扮演重要角色[5]。目前的研究已证实微小RNA(microRNA,miRNA)对GCSCs生物学行为的影响有重要意义,但这些miRNAs对GCSCs的相关调控机制和临床用途仍未完全阐明。因此,更好地了解miRNA在GCSCs中的调控机制,将有助于为胃癌的临床治疗提供新的视角。本综述旨在总结目前miRNA在GCSCs中的调控机制,以及总结基于干性相关基因靶向GCSCs抗肿瘤治疗的措施。
1 miRNA概述
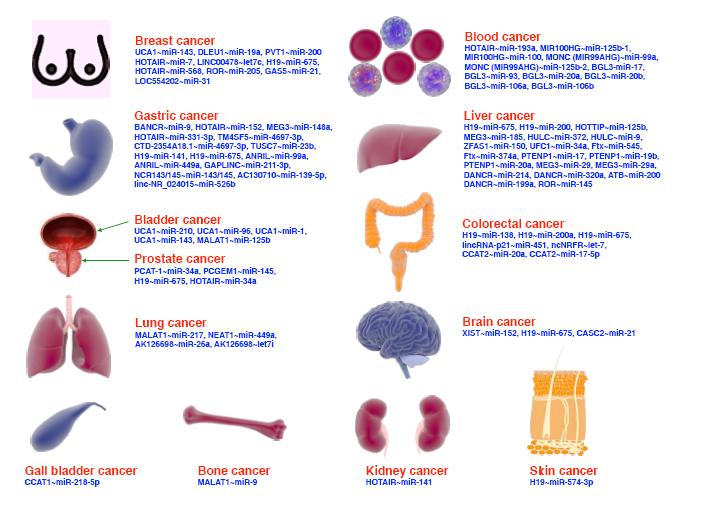
miRNA是一类高度保守的内源性、非编码短链RNA,长度为18~24个核苷酸。通常miRNA与其靶mRNA的3′ -非翻译区(3′ -untranslated region,3′ -UTR)互补靶点结合并相互作用,诱导沉默复合物的形成,引起mRNA的降解或翻译抑制,从而参与一系列重要的细胞过程[6]。到目前为止,miRNA的异常表达已经在包括胃癌在内的多种恶性肿瘤中被发现,其既可以作为抑癌基因发挥作用,如miRNA-195[7]、miRNA-4521[8]、Hsa-miRNA-29c[9]等对胃癌的进展具有抑制作用;也可作为癌基因发挥作用,如miRNA-328-3p[10]、miRNA-135b[11]、miRNA-196a[12]等可以促进胃癌细胞的生长,提高其侵袭能力。诸多研究证实了miRNA潜在的致癌能力,包括靶向基因的降解,而这些基因与正常细胞内稳态的调节有关,如细胞增殖、细胞周期调节、分化、发育、血管生成、细胞凋亡等[13]。
miRNA在调节CSCs的自我更新能力、转移、耐药等方面也发挥着重要作用[14]。值得注意的是,miRNA可以调节与信号通路相关的基因及其产物的表达和功能,通过相关信号通路调节CSCs对环境和生物刺激的反应,在细胞的发生发展过程中起着核心作用。越来越多的研究结果表明,miRNA在确保信号通路的激活、功能发挥等方面起着非常重要的作用[15]。尽管人们在理解miRNA调控CSCs生物学中的作用方面取得了相当大的进展,但仍需进一步研究miRNA的作用机制和临床用途。
2 GCSCs的鉴别与分离
根据CSCs相关理论,肿瘤是由大量异质和分化的癌细胞组成,而这些癌细胞是由少数具有不同增殖和分化能力的CSCs所驱动形成的[16]。CSCs的特点是表现出自我更新能力、高致瘤活性、放化疗抗性以及通过细胞上皮-间充质转化(epithelial-mesenchymal transition,EMT)远处转移的潜能,这使得它们成为与治疗高度相关的特异性靶点[17]。在髓母细胞瘤和甲状腺肿瘤中,CSCs代表肿瘤内的静止细胞群,其特点是对药物治疗的耐药性增加,并且能够驱动肿瘤异质性、复发和转移[18-19]。因此,针对CSCs的研究有助于解决肿瘤复发、转移等临床问题。然而,由于CSCs在实体瘤中的数量极少,因此CSCs的鉴别和分离对于探索其潜在机制和功能至关重要。
研究[20]发现,成球实验是获得CSCs的便捷方法,在添加了生长因子的无血清悬浮培养基中,CSCs可以形成球状细胞并保持自我更新特性,而非CSCs则无法存活。因为球状细胞具有干性特性,例如无限增殖、持续自我更新和CSCs相关基因表达上调。此外,GCSCs中还存在一些特定的分离表面标志物。例如,Takaishi等[21]在胃癌细胞系中发现了相当数量的CD44+ 细胞,当将这些细胞注射到严重免疫缺陷小鼠的胃和皮肤中时,在小鼠体内显示出显著的致瘤性;然而,只有大约5%的CD44+细胞最终被鉴定为GCSCs。因此,单个细胞表面标记很难检测到所有具有GCSCs特征的细胞,仍然存在一些限制和争议。一些研究也证实了结合几种分子作为细胞表面标志物来提高特异性识别和分离GCSCs的能力。例如,CD44+/CD24+[22]、CD133+/CD166+[23]等组合可作为鉴别GCSCs的潜在标志物。CD44不仅可作为鉴别GCSCs的特征性识别靶点,最新的研究还发现CD45–CD44+CD54+ 细胞亚群的数量与肿瘤进展有相关性,可能有助于判断胃癌的分期[24]。除了通过细胞表面标志物分离CSCs外,研究还发现三维肿瘤培养体系可以模拟肿瘤微环境,从而有效地分离CSCs;最新的研究也开始使用这种三维培养体系来分离和培养GCSCs[25-26]。
3 miRNA在GCSCs干性调控中的作用
3.1 miRNA对GCSCs干性调控的双向性
越来越多的研究表明,miRNA可以通过各种靶基因调控GCSCs的干性特征[15]。根据miRNA调控肿瘤发生和发展的作用方式不同,miRNA对胃癌的进展可表现为促进或抑制的双向作用。发挥抑癌基因的作用方面,Hui等[27]研究发现,miRNA-363-3p作为长链非编码核糖核酸(long noncoding ribonucleic acid,lncRNA)FEZ家族锌指1反义RNA1(FEZ family zinc finger 1 antisense RNA 1,FEZF1-AS1)的靶标,因为miRNA-363-3p在GCSCs中的表达被FEZF1-AS1抑制,FEZF1-AS1还可以通过海绵miRNA-363-3p增加高迁移率组AT-hook2(high-mobility group AT-hook 2,HMGA2)的表达;而沉默lncRNA FEZF1-AS1会影响GCSCs的干性特征,并通过miRNA-363-3p介导下调HMGA2,从而抑制GCSCs的体内致瘤能力。此外,还发现miRNA-375可通过miRNA-375/SLC7A11调节轴触发铁死亡,这表明miRNA-375对胃癌干性特征的抑制至关重要[28]。发挥癌基因作用方面,Wu等[29]发现上调miRNA-483-5p可诱导β-catenin及其下游靶分子(包括细胞周期蛋白D1、B淋巴细胞瘤-2和基质金属蛋白酶2)的蛋白表达水平升高,表明miRNA-483-5p通过激活Wnt/β-catenin信号通路调节GCSCs的生长、侵袭和自我更新,从而促进胃癌的进展。Sun等[30]研究发现,应用miRNA-3619-5p模拟物恢复胃癌细胞中的小核仁RNA宿主基因3(small nucleolar RNA host gene 3,SNHG3)表达水平后,八聚体结合转录因子4、性别决定相关基因簇-2和CD44的表达水平在SNHG3过表达后显著增加,这些结果表明,miRNA-3619-5p作为调节轴关键因子促进胃癌细胞的干性特征。总之,在大多数情况下,miRNA可通过以上两种方式共同调控GCSCs的干性特征,从而在胃癌发生发展中具有重要意义。
3.2 miRNA与GCSCs相关信号通路
GCSCs稳态的维持依赖于多条信号通路调控,它们的调控异常可能是GCSCs自我更新失衡的主要原因[31]。如Notch、转化生长因子-β1(trans- forming growth factor-β1,TGF-β1)/Smad、Wnt/β-catenin等信号通路,特定的miRNA通过靶向关键基因,从而参与干性调控的相关信号通路。Xin等[32]研究表明,通过使用甲基化特异性PCR发现,miRNA-7-5p在GCSCs中显著下调是由于启动子区域中DNA 甲基化增加所致,而过表达miRNA-7-5p通过靶向平滑受体蛋白和发状分裂相关增强子1,以及随后在体外抑制Notch和Hedgehog信号通路来减少集落的形成并降低GCSCs的侵袭能力。在体外通过TGF-β1诱导胃癌细胞中的TGF-β信号通路后,观察到miRNA-200a的下调,而EMT相关转录因子E盒结合锌指蛋白1和锌指转录因子则上调,从而获得了CSCs的侵袭和转移特性[33]。除了Notch和TGF-β信号通路外,还发现miRNA的异常表达与Wnt信号通路有关。Song等[34]发现在球状胃癌细胞中溴结构域蛋白4过表达,并通过启动子区域的甲基化来下调miRNA-216a-3p的表达,通过转染miRNA-216a-3p可下调CSCs相关Wnt信号通路的直接靶标Wnt3a的表达。
除了上述信号通路之外,还发现与其他通路相关的miRNA。例如,观察到miRNA-26a在胃癌中表达下调,并通过下调同源盒基因C9(homeobox C9,HOXC9)来调节CSCs相关表面标志物CD44和上皮细胞黏附分子(epithelial cell adhesion molecule,EpCAM)的表达,从而提示HOXC9在体外可以促进胃癌细胞的CSCs表型,由此可发现miRNA-26a的低表达促进了GCSCs的自我更新能力和侵袭能力[35]。最近的研究还发现了miRNA在Hippo途径调控中的作用,Li等[36]报道,miRNA-93-5p通过使Hippo信号通路失活来促进胃癌细胞的进展。而Du等[37]通过生物信息学预测并在胃癌组织和细胞系实验中证实miRNA-95在胃癌中高表达,其通过负向调控双特异性磷酸酶5(dual specificity phosphatase 5,DUSP5)激活丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)通路,并观察到CSCs表面标志物(CD133、CD44、乙醛脱氢酶1和富含亮氨酸重复单位的G蛋白耦联受体5)水平提高。因此,miRNA-95通过DUSP5依赖性MAPK通路调控胃癌进展。
3.3 miRNA与GCSCs转移和耐药
目前胃癌治疗的局限主要是由于其复发转移和对治疗的抵抗,而CSCs的干性特征是导致晚期胃癌患者的不良预后的关键因素。研究[38-39]发现miRNA可以维持CSCs的EMT特性,而获得EMT表型的CSCs不仅可以促进肿瘤侵袭转移,还可以对化疗药物具有抵抗性。Xiao等[40]研究表明,miRNA-711在胃癌中低表达,上调miRNA-711的表达水平抑制了胃癌细胞的侵袭转移及EMT的发生;进一步研究发现,miRNA-711介导的CD44表达下调是通过下调波形蛋白表达和上调E-钙黏蛋白表达实现的,在体内外抑制胃癌细胞的EMT。Pan等[41]通过miRNA微阵列分析显示,在CD44+ 细胞中miRNA-196a-5p的表达显著高于CD44– 细胞,miRNA-196a-5p可通过靶向mRNA的3′ -UTR来调控胃癌组织中Smad4的表达,在GCSCs的EMT和转移侵袭中起到关键作用。此外,Zhang等[42]的研究结果表明,miRNA-345通过靶向表皮生长因子受体激酶底物8使Rac1信号失活,从而抑制细胞转移、EMT和CSCs表型,在胃癌转移中发挥抑制作用。
化疗是继手术和放疗之后恶性肿瘤的主要治疗方案,可用于治疗晚期胃癌患者并延长生存时间。然而,化疗耐药通常会导致治疗失败及患者预后不良。miRNA可以调控GCSCs的化疗药物敏感性,与胃癌复发和转移密切相关。Wu等[43]研究发现DNA甲基化转移酶3B(DNA methyltransferase 3 beta,DNMT3B)作为CSCs的标志物,参与调控CSCs。miRNA-492过表达通过靶向DNMT3B的3′ -UTR、促进CSCs干性特征来刺激肿瘤转移并诱导化疗耐药,而抑制miRNA-492则显著增强胃癌细胞对顺铂化疗的敏感性,并通过靶向DNMT3B抑制胃癌干性特征来抑制胃癌的转移。Shen等[44]通过流式细胞分选技术分离出CD133+/CD44+ 胃癌细胞系MKN45作为 GCSCs进行研究,发现其具有高水平的一切顺势盒2(one cut homeobox 2,ONECUT2)和低水平的hsa-miRNA-15a-5p。进一步体内外实验表明,hsa-miRNA-15a-5p通过靶向ONECUT2调控GCSCs的干性特征、EMT和化疗敏感性。Zhan等[45]发现miRNA-98过表达可抑制干细胞相关基因的表达,抑制GCSCs的自我更新、侵袭转移及致瘤能力;此外,还发现miRNA-98过表达增强了体外顺铂治疗的敏感性;通过异种移植模型,还发现miRNA-98过度表达逆转了紫杉醇对CD44+ GCSCs的耐药性。研究还发现,miRNA还可通过调控细胞凋亡基因的表达导致CSCs耐药现象的发生[46]。在CD44+ 的GCSCs中,miRNA-193a-3p的抑制增加了富含丝氨酸和精氨酸的剪接因子2的表达,并且还改变了多个凋亡基因的水平;抑制miRNA-193a-3p会降低细胞活性并增加凋亡细胞的数量。因此,miRNA-193a-3p可能通过调节线粒体凋亡途径参与顺铂耐药[47]。
最近的研究明确了miRNA在GCSCs中的干性调控机制,通过相关信号通路可以调节GCSCs自我更新或分化能力[15]。此外,miRNA还可通过EMT,在GCSCs的肿瘤转移、侵袭、化疗耐药等方面起着关键的调控作用。miRNA对GCSCs的相关调控的研究可能有助于提高对胃癌的临床治疗效果,通过降低肿瘤转移和复发、预测和逆转肿瘤细胞耐药有着重要的临床意义。
4 miRNA在研发靶向GCSCs治疗方法中的研究进展
胃癌是消化系统常见的恶性肿瘤,由于早期症状不明显,发现时已进展至中晚期,预后较差。虽然目前临床疗效有所提高,但患者的病死率并未得到有效降低[48]。研究证实,胃癌的治疗进展主要集中于新的治疗靶点的确定[3]。Wei等[49]通过一系列机器学习方法为胃癌开发了胃癌干细胞相关评分(GCSC-related score,GCScore),基于GCScore研究GCSCs的免疫学特征和潜在的免疫逃逸机制,提出GCScore可以作为预后和免疫治疗反应的预测因子,确定高GCScore胃癌患者的4个潜在靶点(DARS2、SLC2A1、TPMT和ATPV1C1),为GCSCs的精准治疗提供新策略。而miRNA作为重要的基因表达调控因子,除了在调节GCSCs干性特征中发挥着关键作用之外,通过上述诸多调控机制,其中一些miRNA在研发靶向GCSCs的治疗方法中起着重要作用。因此,在肿瘤治疗过程中通过沉默或过表达调节特定的miRNA是一个重要的探索途径。
某些miRNA可促进GCSCs对化疗药物的耐药性,通过沉默这些miRNA可以提高胃癌患者对治疗的敏感性。例如,抑制GCSCs中miRNA-193-3p的表达会使CD44+ 的GCSCs对顺铂敏感性提高[47]。除了通过沉默某些可以促进GCSCs干性特征的miRNA以提高对化疗药物的敏感性外,在对化疗药物耐药的GCSCs中还发现一些miRNA的表达下调,与沉默促进化疗耐药的miRNA治疗相似,重新恢复这些被下调的miRNA的表达量能提高对治疗的敏感度,可能也是一种有前景的治疗方式。有研究[50]发现用miRNA模拟miR-876-3p治疗可增强GCSCs对顺铂的敏感性。此外,中药成分益气补肾含药血清可通过上调GCSCs中miRNA-139-5p的表达水平,作用其靶基因Notch1,通过调控miRNA-139-5p/Notch1/Jagged1信号通路发挥抗胃癌增殖转移的作用[51]。一种miRNA-34模拟物MRX34被开发用于肿瘤治疗,并进行了Ⅰ期临床试验。由于可能与miRNA-34模拟物传递载体相关的严重免疫反应,该试验被终止,但对基于miRNA的表达靶向CSCs从而治疗胃癌,仍然做出了重要贡献[52-53]。
基于miRNA的疗法,无论是恢复还是抑制miRNA的表达和活性,都具有很大的临床应用前景。然而,将miRNA有效递送至靶组织是miRNA治疗向临床过渡的主要挑战。利用与抗体和(或)多肽结合的纳米颗粒高效持续靶向释放miRNA/抗miRNA,可以减少所需的治疗剂量,同时将细胞毒性降至最低[54]。Liu等[55]通过凝胶刺激纳米颗粒来共同递送miRNA-200c和多西紫杉醇(docetaxel,DOC),比较miRNA-200c/DOC纳米颗粒与DOC纳米颗粒在移植瘤小鼠中的抗肿瘤作用,发现miRNA-200c/DOC纳米颗粒表现出较高的miRNA-200c转染效率,显著上调miRNA-200c的表达,并在较长一段时间内降低β-微管蛋白Ⅲ的表达,显著增强DOC的细胞毒性,从而产生协同的肿瘤抑制作用;miRNA-200c/DOC纳米颗粒还通过下调CSCs表面标志物CD44的表达抑制GCSCs;此外,当将miRNA-200c/DOC纳米颗粒治疗后的残余肿瘤再次移植到裸鼠体内时,肿瘤的生长速度明显减慢。因此,miRNA-200c/DOC纳米颗粒可能为miRNA和化疗药物的联合运用提供一种很有前途的方式,同时抑制CSCs和非CSCs。最近的研究[56]还发现,miRNA-200c纳米颗粒联合放疗对胃癌细胞具有协同抑制作用。可能的机制包括:通过纳米颗粒将miRNA-200c稳定有效地递送到胃癌细胞中以及通过抑制TGF-β1和CD44来逆转EMT。因此,miRNA-200c纳米颗粒联合放疗可能是治疗胃癌的潜在策略。
5 小结与展望
GCSCs已被研究发现是胃癌治疗的主要障碍,因为它们在肿瘤转移、复发以及对治疗的抵抗中发挥重要作用,制定能够解决这一障碍的治疗策略非常重要。在本篇综述中,回顾了miRNA的异常表达对GCSCs干性特征的调控以及基于miRNA靶向GCSCs抗肿瘤治疗的最新进展。更好地了解miRNA的分子机制及其参与GCSCs调控的基因靶点可以为GCSCs的靶向治疗开辟新的策略。虽然用于介导传递miRNA的纳米颗粒可用于模拟miRNA发挥其抑癌基因的作用,从而抑制GCSCs的干性特征,但基于miRNA药物的安全性和有效性仍然是值得关注的问题,需要更多研究来优化miRNA由基础实验向临床应用的转化。因此,以miRNA为调控靶点预防及克服GCSCs转移、耐药等干性特征对改善胃癌的治疗是值得探索研究的重要方向。
重要声明
利益冲突声明:本文全体作者阅读并理解了《中国普外基础与临床杂志》的政策声明,我们无相互竞争的利益。
作者贡献声明:林浩主要撰写论文;王朝樣、张海宝、巨家华和俞永江对论文进行指导并修改。
胃癌是严重威胁人类健康的恶性肿瘤之一,全球每年新发病例约93万例,近70万人死亡,其发病率和病死率分别位居所有恶性肿瘤的第4位和第2位[1]。胃镜等临床诊断技术的进步提高了胃癌患者的早期检出率[2]。对于早期胃癌患者,手术切除有着良好的疗效。然而,对于晚期患者,由于存在局部浸润和远处转移,治疗选择仍然有限,手术切除效果不理想。进一步鉴定调控胃癌进展的关键分子可为晚期胃癌患者提供新的治疗靶点[3]。肿瘤干细胞(cancer stem cells,CSCs)是能够引起肿瘤生长并具有自我更新、多向分化能力的一类细胞亚群,可以分化成大量新的肿瘤细胞,与肿瘤的恶性生物学行为及不良预后有着重要的关系[4]。研究发现胃癌干细胞(gastric cancer stem cells,GCSCs)在胃癌疾病进展过程中扮演重要角色[5]。目前的研究已证实微小RNA(microRNA,miRNA)对GCSCs生物学行为的影响有重要意义,但这些miRNAs对GCSCs的相关调控机制和临床用途仍未完全阐明。因此,更好地了解miRNA在GCSCs中的调控机制,将有助于为胃癌的临床治疗提供新的视角。本综述旨在总结目前miRNA在GCSCs中的调控机制,以及总结基于干性相关基因靶向GCSCs抗肿瘤治疗的措施。
1 miRNA概述
miRNA是一类高度保守的内源性、非编码短链RNA,长度为18~24个核苷酸。通常miRNA与其靶mRNA的3′ -非翻译区(3′ -untranslated region,3′ -UTR)互补靶点结合并相互作用,诱导沉默复合物的形成,引起mRNA的降解或翻译抑制,从而参与一系列重要的细胞过程[6]。到目前为止,miRNA的异常表达已经在包括胃癌在内的多种恶性肿瘤中被发现,其既可以作为抑癌基因发挥作用,如miRNA-195[7]、miRNA-4521[8]、Hsa-miRNA-29c[9]等对胃癌的进展具有抑制作用;也可作为癌基因发挥作用,如miRNA-328-3p[10]、miRNA-135b[11]、miRNA-196a[12]等可以促进胃癌细胞的生长,提高其侵袭能力。诸多研究证实了miRNA潜在的致癌能力,包括靶向基因的降解,而这些基因与正常细胞内稳态的调节有关,如细胞增殖、细胞周期调节、分化、发育、血管生成、细胞凋亡等[13]。
miRNA在调节CSCs的自我更新能力、转移、耐药等方面也发挥着重要作用[14]。值得注意的是,miRNA可以调节与信号通路相关的基因及其产物的表达和功能,通过相关信号通路调节CSCs对环境和生物刺激的反应,在细胞的发生发展过程中起着核心作用。越来越多的研究结果表明,miRNA在确保信号通路的激活、功能发挥等方面起着非常重要的作用[15]。尽管人们在理解miRNA调控CSCs生物学中的作用方面取得了相当大的进展,但仍需进一步研究miRNA的作用机制和临床用途。
2 GCSCs的鉴别与分离
根据CSCs相关理论,肿瘤是由大量异质和分化的癌细胞组成,而这些癌细胞是由少数具有不同增殖和分化能力的CSCs所驱动形成的[16]。CSCs的特点是表现出自我更新能力、高致瘤活性、放化疗抗性以及通过细胞上皮-间充质转化(epithelial-mesenchymal transition,EMT)远处转移的潜能,这使得它们成为与治疗高度相关的特异性靶点[17]。在髓母细胞瘤和甲状腺肿瘤中,CSCs代表肿瘤内的静止细胞群,其特点是对药物治疗的耐药性增加,并且能够驱动肿瘤异质性、复发和转移[18-19]。因此,针对CSCs的研究有助于解决肿瘤复发、转移等临床问题。然而,由于CSCs在实体瘤中的数量极少,因此CSCs的鉴别和分离对于探索其潜在机制和功能至关重要。
研究[20]发现,成球实验是获得CSCs的便捷方法,在添加了生长因子的无血清悬浮培养基中,CSCs可以形成球状细胞并保持自我更新特性,而非CSCs则无法存活。因为球状细胞具有干性特性,例如无限增殖、持续自我更新和CSCs相关基因表达上调。此外,GCSCs中还存在一些特定的分离表面标志物。例如,Takaishi等[21]在胃癌细胞系中发现了相当数量的CD44+ 细胞,当将这些细胞注射到严重免疫缺陷小鼠的胃和皮肤中时,在小鼠体内显示出显著的致瘤性;然而,只有大约5%的CD44+细胞最终被鉴定为GCSCs。因此,单个细胞表面标记很难检测到所有具有GCSCs特征的细胞,仍然存在一些限制和争议。一些研究也证实了结合几种分子作为细胞表面标志物来提高特异性识别和分离GCSCs的能力。例如,CD44+/CD24+[22]、CD133+/CD166+[23]等组合可作为鉴别GCSCs的潜在标志物。CD44不仅可作为鉴别GCSCs的特征性识别靶点,最新的研究还发现CD45–CD44+CD54+ 细胞亚群的数量与肿瘤进展有相关性,可能有助于判断胃癌的分期[24]。除了通过细胞表面标志物分离CSCs外,研究还发现三维肿瘤培养体系可以模拟肿瘤微环境,从而有效地分离CSCs;最新的研究也开始使用这种三维培养体系来分离和培养GCSCs[25-26]。
3 miRNA在GCSCs干性调控中的作用
3.1 miRNA对GCSCs干性调控的双向性
越来越多的研究表明,miRNA可以通过各种靶基因调控GCSCs的干性特征[15]。根据miRNA调控肿瘤发生和发展的作用方式不同,miRNA对胃癌的进展可表现为促进或抑制的双向作用。发挥抑癌基因的作用方面,Hui等[27]研究发现,miRNA-363-3p作为长链非编码核糖核酸(long noncoding ribonucleic acid,lncRNA)FEZ家族锌指1反义RNA1(FEZ family zinc finger 1 antisense RNA 1,FEZF1-AS1)的靶标,因为miRNA-363-3p在GCSCs中的表达被FEZF1-AS1抑制,FEZF1-AS1还可以通过海绵miRNA-363-3p增加高迁移率组AT-hook2(high-mobility group AT-hook 2,HMGA2)的表达;而沉默lncRNA FEZF1-AS1会影响GCSCs的干性特征,并通过miRNA-363-3p介导下调HMGA2,从而抑制GCSCs的体内致瘤能力。此外,还发现miRNA-375可通过miRNA-375/SLC7A11调节轴触发铁死亡,这表明miRNA-375对胃癌干性特征的抑制至关重要[28]。发挥癌基因作用方面,Wu等[29]发现上调miRNA-483-5p可诱导β-catenin及其下游靶分子(包括细胞周期蛋白D1、B淋巴细胞瘤-2和基质金属蛋白酶2)的蛋白表达水平升高,表明miRNA-483-5p通过激活Wnt/β-catenin信号通路调节GCSCs的生长、侵袭和自我更新,从而促进胃癌的进展。Sun等[30]研究发现,应用miRNA-3619-5p模拟物恢复胃癌细胞中的小核仁RNA宿主基因3(small nucleolar RNA host gene 3,SNHG3)表达水平后,八聚体结合转录因子4、性别决定相关基因簇-2和CD44的表达水平在SNHG3过表达后显著增加,这些结果表明,miRNA-3619-5p作为调节轴关键因子促进胃癌细胞的干性特征。总之,在大多数情况下,miRNA可通过以上两种方式共同调控GCSCs的干性特征,从而在胃癌发生发展中具有重要意义。
3.2 miRNA与GCSCs相关信号通路
GCSCs稳态的维持依赖于多条信号通路调控,它们的调控异常可能是GCSCs自我更新失衡的主要原因[31]。如Notch、转化生长因子-β1(trans- forming growth factor-β1,TGF-β1)/Smad、Wnt/β-catenin等信号通路,特定的miRNA通过靶向关键基因,从而参与干性调控的相关信号通路。Xin等[32]研究表明,通过使用甲基化特异性PCR发现,miRNA-7-5p在GCSCs中显著下调是由于启动子区域中DNA 甲基化增加所致,而过表达miRNA-7-5p通过靶向平滑受体蛋白和发状分裂相关增强子1,以及随后在体外抑制Notch和Hedgehog信号通路来减少集落的形成并降低GCSCs的侵袭能力。在体外通过TGF-β1诱导胃癌细胞中的TGF-β信号通路后,观察到miRNA-200a的下调,而EMT相关转录因子E盒结合锌指蛋白1和锌指转录因子则上调,从而获得了CSCs的侵袭和转移特性[33]。除了Notch和TGF-β信号通路外,还发现miRNA的异常表达与Wnt信号通路有关。Song等[34]发现在球状胃癌细胞中溴结构域蛋白4过表达,并通过启动子区域的甲基化来下调miRNA-216a-3p的表达,通过转染miRNA-216a-3p可下调CSCs相关Wnt信号通路的直接靶标Wnt3a的表达。
除了上述信号通路之外,还发现与其他通路相关的miRNA。例如,观察到miRNA-26a在胃癌中表达下调,并通过下调同源盒基因C9(homeobox C9,HOXC9)来调节CSCs相关表面标志物CD44和上皮细胞黏附分子(epithelial cell adhesion molecule,EpCAM)的表达,从而提示HOXC9在体外可以促进胃癌细胞的CSCs表型,由此可发现miRNA-26a的低表达促进了GCSCs的自我更新能力和侵袭能力[35]。最近的研究还发现了miRNA在Hippo途径调控中的作用,Li等[36]报道,miRNA-93-5p通过使Hippo信号通路失活来促进胃癌细胞的进展。而Du等[37]通过生物信息学预测并在胃癌组织和细胞系实验中证实miRNA-95在胃癌中高表达,其通过负向调控双特异性磷酸酶5(dual specificity phosphatase 5,DUSP5)激活丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)通路,并观察到CSCs表面标志物(CD133、CD44、乙醛脱氢酶1和富含亮氨酸重复单位的G蛋白耦联受体5)水平提高。因此,miRNA-95通过DUSP5依赖性MAPK通路调控胃癌进展。
3.3 miRNA与GCSCs转移和耐药
目前胃癌治疗的局限主要是由于其复发转移和对治疗的抵抗,而CSCs的干性特征是导致晚期胃癌患者的不良预后的关键因素。研究[38-39]发现miRNA可以维持CSCs的EMT特性,而获得EMT表型的CSCs不仅可以促进肿瘤侵袭转移,还可以对化疗药物具有抵抗性。Xiao等[40]研究表明,miRNA-711在胃癌中低表达,上调miRNA-711的表达水平抑制了胃癌细胞的侵袭转移及EMT的发生;进一步研究发现,miRNA-711介导的CD44表达下调是通过下调波形蛋白表达和上调E-钙黏蛋白表达实现的,在体内外抑制胃癌细胞的EMT。Pan等[41]通过miRNA微阵列分析显示,在CD44+ 细胞中miRNA-196a-5p的表达显著高于CD44– 细胞,miRNA-196a-5p可通过靶向mRNA的3′ -UTR来调控胃癌组织中Smad4的表达,在GCSCs的EMT和转移侵袭中起到关键作用。此外,Zhang等[42]的研究结果表明,miRNA-345通过靶向表皮生长因子受体激酶底物8使Rac1信号失活,从而抑制细胞转移、EMT和CSCs表型,在胃癌转移中发挥抑制作用。
化疗是继手术和放疗之后恶性肿瘤的主要治疗方案,可用于治疗晚期胃癌患者并延长生存时间。然而,化疗耐药通常会导致治疗失败及患者预后不良。miRNA可以调控GCSCs的化疗药物敏感性,与胃癌复发和转移密切相关。Wu等[43]研究发现DNA甲基化转移酶3B(DNA methyltransferase 3 beta,DNMT3B)作为CSCs的标志物,参与调控CSCs。miRNA-492过表达通过靶向DNMT3B的3′ -UTR、促进CSCs干性特征来刺激肿瘤转移并诱导化疗耐药,而抑制miRNA-492则显著增强胃癌细胞对顺铂化疗的敏感性,并通过靶向DNMT3B抑制胃癌干性特征来抑制胃癌的转移。Shen等[44]通过流式细胞分选技术分离出CD133+/CD44+ 胃癌细胞系MKN45作为 GCSCs进行研究,发现其具有高水平的一切顺势盒2(one cut homeobox 2,ONECUT2)和低水平的hsa-miRNA-15a-5p。进一步体内外实验表明,hsa-miRNA-15a-5p通过靶向ONECUT2调控GCSCs的干性特征、EMT和化疗敏感性。Zhan等[45]发现miRNA-98过表达可抑制干细胞相关基因的表达,抑制GCSCs的自我更新、侵袭转移及致瘤能力;此外,还发现miRNA-98过表达增强了体外顺铂治疗的敏感性;通过异种移植模型,还发现miRNA-98过度表达逆转了紫杉醇对CD44+ GCSCs的耐药性。研究还发现,miRNA还可通过调控细胞凋亡基因的表达导致CSCs耐药现象的发生[46]。在CD44+ 的GCSCs中,miRNA-193a-3p的抑制增加了富含丝氨酸和精氨酸的剪接因子2的表达,并且还改变了多个凋亡基因的水平;抑制miRNA-193a-3p会降低细胞活性并增加凋亡细胞的数量。因此,miRNA-193a-3p可能通过调节线粒体凋亡途径参与顺铂耐药[47]。
最近的研究明确了miRNA在GCSCs中的干性调控机制,通过相关信号通路可以调节GCSCs自我更新或分化能力[15]。此外,miRNA还可通过EMT,在GCSCs的肿瘤转移、侵袭、化疗耐药等方面起着关键的调控作用。miRNA对GCSCs的相关调控的研究可能有助于提高对胃癌的临床治疗效果,通过降低肿瘤转移和复发、预测和逆转肿瘤细胞耐药有着重要的临床意义。
4 miRNA在研发靶向GCSCs治疗方法中的研究进展
胃癌是消化系统常见的恶性肿瘤,由于早期症状不明显,发现时已进展至中晚期,预后较差。虽然目前临床疗效有所提高,但患者的病死率并未得到有效降低[48]。研究证实,胃癌的治疗进展主要集中于新的治疗靶点的确定[3]。Wei等[49]通过一系列机器学习方法为胃癌开发了胃癌干细胞相关评分(GCSC-related score,GCScore),基于GCScore研究GCSCs的免疫学特征和潜在的免疫逃逸机制,提出GCScore可以作为预后和免疫治疗反应的预测因子,确定高GCScore胃癌患者的4个潜在靶点(DARS2、SLC2A1、TPMT和ATPV1C1),为GCSCs的精准治疗提供新策略。而miRNA作为重要的基因表达调控因子,除了在调节GCSCs干性特征中发挥着关键作用之外,通过上述诸多调控机制,其中一些miRNA在研发靶向GCSCs的治疗方法中起着重要作用。因此,在肿瘤治疗过程中通过沉默或过表达调节特定的miRNA是一个重要的探索途径。
某些miRNA可促进GCSCs对化疗药物的耐药性,通过沉默这些miRNA可以提高胃癌患者对治疗的敏感性。例如,抑制GCSCs中miRNA-193-3p的表达会使CD44+ 的GCSCs对顺铂敏感性提高[47]。除了通过沉默某些可以促进GCSCs干性特征的miRNA以提高对化疗药物的敏感性外,在对化疗药物耐药的GCSCs中还发现一些miRNA的表达下调,与沉默促进化疗耐药的miRNA治疗相似,重新恢复这些被下调的miRNA的表达量能提高对治疗的敏感度,可能也是一种有前景的治疗方式。有研究[50]发现用miRNA模拟miR-876-3p治疗可增强GCSCs对顺铂的敏感性。此外,中药成分益气补肾含药血清可通过上调GCSCs中miRNA-139-5p的表达水平,作用其靶基因Notch1,通过调控miRNA-139-5p/Notch1/Jagged1信号通路发挥抗胃癌增殖转移的作用[51]。一种miRNA-34模拟物MRX34被开发用于肿瘤治疗,并进行了Ⅰ期临床试验。由于可能与miRNA-34模拟物传递载体相关的严重免疫反应,该试验被终止,但对基于miRNA的表达靶向CSCs从而治疗胃癌,仍然做出了重要贡献[52-53]。
基于miRNA的疗法,无论是恢复还是抑制miRNA的表达和活性,都具有很大的临床应用前景。然而,将miRNA有效递送至靶组织是miRNA治疗向临床过渡的主要挑战。利用与抗体和(或)多肽结合的纳米颗粒高效持续靶向释放miRNA/抗miRNA,可以减少所需的治疗剂量,同时将细胞毒性降至最低[54]。Liu等[55]通过凝胶刺激纳米颗粒来共同递送miRNA-200c和多西紫杉醇(docetaxel,DOC),比较miRNA-200c/DOC纳米颗粒与DOC纳米颗粒在移植瘤小鼠中的抗肿瘤作用,发现miRNA-200c/DOC纳米颗粒表现出较高的miRNA-200c转染效率,显著上调miRNA-200c的表达,并在较长一段时间内降低β-微管蛋白Ⅲ的表达,显著增强DOC的细胞毒性,从而产生协同的肿瘤抑制作用;miRNA-200c/DOC纳米颗粒还通过下调CSCs表面标志物CD44的表达抑制GCSCs;此外,当将miRNA-200c/DOC纳米颗粒治疗后的残余肿瘤再次移植到裸鼠体内时,肿瘤的生长速度明显减慢。因此,miRNA-200c/DOC纳米颗粒可能为miRNA和化疗药物的联合运用提供一种很有前途的方式,同时抑制CSCs和非CSCs。最近的研究[56]还发现,miRNA-200c纳米颗粒联合放疗对胃癌细胞具有协同抑制作用。可能的机制包括:通过纳米颗粒将miRNA-200c稳定有效地递送到胃癌细胞中以及通过抑制TGF-β1和CD44来逆转EMT。因此,miRNA-200c纳米颗粒联合放疗可能是治疗胃癌的潜在策略。
5 小结与展望
GCSCs已被研究发现是胃癌治疗的主要障碍,因为它们在肿瘤转移、复发以及对治疗的抵抗中发挥重要作用,制定能够解决这一障碍的治疗策略非常重要。在本篇综述中,回顾了miRNA的异常表达对GCSCs干性特征的调控以及基于miRNA靶向GCSCs抗肿瘤治疗的最新进展。更好地了解miRNA的分子机制及其参与GCSCs调控的基因靶点可以为GCSCs的靶向治疗开辟新的策略。虽然用于介导传递miRNA的纳米颗粒可用于模拟miRNA发挥其抑癌基因的作用,从而抑制GCSCs的干性特征,但基于miRNA药物的安全性和有效性仍然是值得关注的问题,需要更多研究来优化miRNA由基础实验向临床应用的转化。因此,以miRNA为调控靶点预防及克服GCSCs转移、耐药等干性特征对改善胃癌的治疗是值得探索研究的重要方向。
重要声明
利益冲突声明:本文全体作者阅读并理解了《中国普外基础与临床杂志》的政策声明,我们无相互竞争的利益。
作者贡献声明:林浩主要撰写论文;王朝樣、张海宝、巨家华和俞永江对论文进行指导并修改。