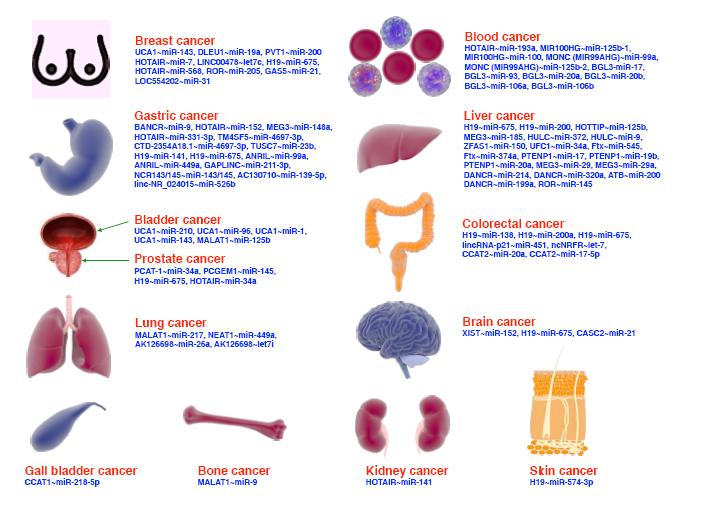

环状 RNA(circular RNA,circRNA)是一类新型的非编码 RNA,以共价键形成闭合环形结构,较线性 RNA 更稳定。CircRNA 作为在真核生物中的高丰度 RNA,参与调控靶基因表达,与多种疾病有着密切联系,有作为临床诊断及预后标志物的潜在应用价值。该文概述了 circRNA 分子的分类、生物学功能(包括作为微 RNA 分子海绵、调控基因转录、调节 RNA 结合蛋白以及其潜在的翻译功能),并就近年来 circRNA 在类风湿关节炎中的最新研究进展作一综述。
引用本文: 陈一丹, 林桑, 刘欢, 尹耕. 环状 RNA 在类风湿关节炎中的研究进展. 华西医学, 2019, 34(6): 693-697. doi: 10.7507/1002-0179.201904058 复制
环状 RNA(circular RNA,circRNA)是由前体 RNA 可变剪接产生的一类不具有游离的 5’ 端帽子和 3’ 端 poly(A)尾巴、以共价键形成环形结构的非编码 RNA 分子,且不易被核酸外切酶 RNaseR 降解[1]。1976 年人们在仙台病毒(Sendai virus)中首次发现 circRNA[2]。早期研究认为 circRNA 是一类剪接副产物或因错误剪接而形成的低丰度 RNA 分子[3],未引起关注。随着测序技术及生物信息学的发展和广泛应用,大量的 circRNA 被发现,如 Jeck 等[4]在人类成纤维细胞中检测出超过 25 000 种 circRNA;Salzman 等[5]发现了大量与人类基因表达密切相关的 circRNA。研究表明 circRNA 具有存在广泛、高度稳定、时序及疾病特异性等生物学特征,对于基因表达也起到调控作用[5-7]。
类风湿关节炎(rheumatoid arthritis,RA)是一种以慢性滑膜炎为特征的自身免疫性疾病,其发病受多因素影响,最终可能出现不同程度关节软骨和骨质破坏[8-9]。现今越来越多的报道揭示了 circRNA 与多种人类疾病有关,如血管动脉粥样硬化性疾病、癌症和自身免疫性疾病等[10-14],预示着 circRNA 可能成为新一代的生物标志物或治疗靶点。而到目前为止,还很少有研究探讨 circRNA 与 RA 之间的关系,因此本文就 circRNA 在 RA 中的最新研究进展作一综述。
1 CircRNA 的分类
目前,根据形成方式的不同,内源性 circRNA 主要分为 3 种:外显子 circRNA(exonic circRNA,ecircRNA)、内含子 circRNA(circular intronic RNA,ciRNA)以及外显子-内含子 circRNA(exonic-intronic circRNA,EI ciRNA)[15]。这些 circRNA 分子间存在竞争性平衡关系,可影响 mRNA 的表达。
1.1 ecircRNA
ecircRNA 为由外显子序列构成的 circRNA,主要存在于细胞质中[4],也是目前研究得最多的 circRNA。CircRNA 的外显子环化包括套索驱动环化和内含子配对驱动环化两种模式。前者是由前体 mRNA 在剪接过程中发生部分折叠而拉近非相邻的外显子,发生外显子跳跃的经典剪接。被跳过外显子的上、下游结合形成套索前体,随后内部剪接去除内含子,形成 ecircRNA[16]。后者是两侧内含子的反向互补序列碱基配对,诱导二者间下游剪接供体与上游剪接受体结合,而环化的外显子被释放成为了 circRNA 分子[17]。
1.2 ciRNA
内含子序列占人类基因组的 20% 以上,但人们普遍认为大多数内含子或内含子片段是不稳定的[18]。Zhang 等[19]提出了内含子来源的非编码 circRNA。该研究也提示了非编码内含子转录子在其亲本编码基因上的顺式调控作用。机体内置于细胞核中的 ciRNA 是由内含子独立环化形成,并主要源于内含子的自我剪接。ciRNA 的形成有赖于含有 3’ 剪接位点附近的 11 nt C 富集区域和邻近 5’ 端的 7 nt GU 富集元件,具有少量的微 RNA(microRNA,miRNA)靶点[20]。上述特殊结构使内含子在剪接过程中避免被脱支酶消化而形成 ciRNA[6]。
1.3 EI ciRNA
EI ciRNA 主要定位于细胞核。在 circRNA 结构中环化的外显子中间保留了内含子序列,我们称之为外显子-内含子环状结构或 EI ciRNA[21],但具体发生机制尚不明确。Li 等[18]研究发现 EI ciRNA 可通过 U1 核小 RNA 和 EI ciRNA 之间的特定的 RNA-RNA 相互作用进行转录控制的调控策略,增强亲本基因的表达。上述研究揭示了基因表达在转录及转录后水平的多样性和复杂性。
2 CircRNA 的生物学功能
2.1 MiRNA 分子海绵
CircRNA 富含与 miRNA 的结合位点,可抑制 mRNA 与靶基因相结合,从而发挥 miRNA“海绵”效应,这是目前 circRNA 被报道最多的功能。Hansen 等[22]研究发现,小脑变性相关蛋白 1 反义转录物(antisense to the cerebellar degeneration-related protein 1 transcript,CDR1as)是小脑变性相关蛋白 1(cerebellar degeneration related protein 1,CDR1)基因的环状天然反义转录物,也被称为 miRNA-7(miR-7)的 circRNA 海绵(ciRS-7),对 miR-7 起到负调控作用[23],上调其靶基因的表达水平,提示 circRNA 可通过参与到竞争的内源 RNA 网络,从而在转录水平上对靶基因进行调控来发挥生物学作用[24]。在胚胎中超高表达 CDR1as 或沉默 miR-7 可导致斑马鱼中脑缩小[25],暗示了 CDR1as 可作为 miR-7 分子海绵的作用。Bai 等[26]研究发现 circDLGAP4 可作为内源性 miRNA-143(miR-143)海绵来抑制其活性,解除 miR-143 对 HECTD1 基因表达的抑制作用,可见,circRNA 可提高靶基因的表达水平[27],当 miRNA 被 circRNA 吸附时,mRNA 翻译合成蛋白质[28]。虽然当前,circRNA 起 miRNA 的海绵作用还在肿瘤[29]、卵泡发育[30]及骨肉瘤的生长[31]等研究中也有体现,但 circRNA 是否普遍具有 miRNA 海绵作用还有待进一步验证。
2.2 CircRNA 调控基因转录
与传统线性 RNA 不同,circRNA 在真核转录组中大量存在。一些内含子来源的 circRNA 存在于细胞核中,可与 RNA 聚合酶Ⅱ转录复合体结合,以顺式作用方式对亲本基因的转录进行调控[11],进而影响靶基因表达。如 ci-ankrd52 是来源于锚蛋白重复结构域 52(ankyrin repeat domain 52,ANKRD52)基因第 2 个内含子的 circRNA,研究表明,它可参与 RNA 聚合酶Ⅱ的延伸机制,起正向调控基因的转录活性作用,而敲除 ci-ankrd52 后,ankrd52 mRNA 表达显著减少,circ-sirt7 也具有与 ci-ankrd52 相同的特征[19],表明细胞核内的 circRNA 可能参与基因的转录调控。此外,还有一些定位在细胞核中的外显子与内含子共同来源的 EI ciRNA(如 circ-EIF3J 和 circ-PAIP2),可与小核糖体 U1 snRNP 或 RNA 聚合酶Ⅱ相互作用促进亲本基因的转录[18],也暗示了 circRNA 的种类及功能具有多样性。
2.3 CircRNA 调节 RNA 结合蛋白
此前已有研究报道线性 RNA 可吸附蛋白因子[32],现研究发现 circRNA 也能与有关蛋白[主要是 RNA 结合蛋白(RNA-binding protein,RBP)]结合形成复合体,进而发挥生物学功能[17]。例如,circRNA(如 CDR1as 和 Sry)可以与 Argonaute(AGO)蛋白结合,调控 AGO mRNA 的翻译过程[33]。Ashwal-Fluss 等[16]发现作为 RBP 的剪接因子肌盲蛋白(muscle blind-like,MBL)可结合其亲本基因的外显子并促进其环化形成 circMBL,二者相互作用可降低 MBL 有效浓度,减少 circMBL 生成。
2.4 CircRNA 具有潜在翻译功能
一般情况下,很少有 circRNA 被翻译成蛋白质。但 Chen 等[34]报道了 circRNA 可启动并编码蛋白。之后 Perriman 等[35]研究出包含一个简单绿色荧光蛋白开放阅读框的 circRNA,它可以在大肠杆菌中指导绿色荧光蛋白的表达。Abe 等[36]验证了在无细胞大肠杆菌翻译系统和人类细胞中,circRNA 可通过滚动循环扩增途径有效地被翻译,并产生丰富的蛋白质产物,也表明这些在人类细胞中大量存在的 ecircRNA 所具有较大的翻译潜能。依赖 N6-甲基腺甲基化修饰[37]和依赖内部核糖体嵌入位点[38]是目前报道最多的 circRNA 两种可能的翻译机制。但 circRNA 在翻译控制中的作用仍然值得进一步研究,且目前为止尚无真核生物细胞自然产生此类具有翻译功能 circRNA 的证据。
3 CircRNA 与 RA
近些年测序结果表明 circRNA 在 RA 患者及健康人外周血单核细胞中存在表达异常,表 1 总结了与 RA 有关的 circRNA[39-45]。
3.1 高表达 circRNA
Ouyang 等[40]通过微阵列芯片技术筛选出差异表达的 circRNA 后使用实时荧光定量聚合酶链反应(real-time quantitative polymerase chain reaction,qRT-PCR)验证了 30 例 RA 患者及 25 例健康人的外周血单核细胞,发现与健康对照组相比,RA 患者中的 circRNA_092516、circRNA_003524、circRNA_103047、circRNA_104871 及 circRNA_101873 表达显著上调,其中受试者工作特征(receiver operating characteristic,ROC)曲线分析提示 circRNA_104871 对于 RA 有较高的诊断价值[ROC 曲线下面积(area under curve,AUC)=0.833,95% 置信区间(0.721,0.944),P<0.001]。进一步研究发现这些 RA 患者外周血单核细胞中具有显著差异性表达的 circRNA 与 28 处关节疾病活动度评分(Disease Activity Score 28,DAS28)、C 反应蛋白、红细胞沉降率、健康评估问卷(Health Assessment Questionnaire,HAQ)均无相关性,表明这些 circRNA 可能不是疾病活动度或者全身炎症反应的相关生物标志物。Zheng 等[41]通过微阵列芯片检测 10 例 RA 患者及 10 例健康对照者外周血单核细胞中的 cricRNA 表达情况,发现有 255 个 circRNA 表达显著上调,329 种 cricRNA 显著下调,并采用 qRT-PCR 验证了 hsa_circ_0092285、hsa_circ_ 0058794 表达显著上调。而 hsa_circ_0092285 可能通过与 PNKP 基因作用参与到 RA 的氧化应激与炎症反应中。hsa_circ_0058794 由 AGAP1 基因拼接而来,进一步研究发现它能促进滑膜细胞侵袭软骨组织[46],有可能成为 RA 患者预后评估的生物标志物。Tang 等[42]报道 ciRS-7 在 RA 患者外周血中显著高表达,并发现 RA 患者 ciRS-7 的相对表达量与血浆抗环瓜氨酸多肽抗体水平呈正相关(r=0.564 0,P=0.014 8);该研究进一步揭示哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)mRNA 在 RA 患者外周血中的表达显著高于健康对照组,并与 ciRS-7 表达呈正相关,与 miR-7 表达呈负相关,推测在 RA 患者中 ciRS-7 可能作为 miR-7 的“海绵”并“吸收”一定量的 miR-7 进而上调 mTOR 的表达。这些结果表明 ciRS-7 可能是 RA 潜在的生物标志物(AUC=0.766),为 RA 的诊断和靶向治疗提供了新的思路。
3.2 低表达 circRNA
有研究发现源自于类固醇脱氢酶样蛋白 2(hydroxysteroid dehydrogenase like 2,HSDL2)基因的 hsa_circ_0088088 表达水平在 RA 患者外周血中明显下调[41],并可能增加 RA 患者心血管事件的风险[47]。就机制而言,HSDL2 参与了脂肪酸和胆固醇的代谢和稳态的定位[48],其下调可能影响脂质分解并增加游离脂肪酸水平[49],从而增加 RA 患者动脉粥样硬化的风险[50]。与健康人相比,在 RA 患者外周血中显著下调的 hsa_circ_0038644 于 PRKCB 中编码[41],而 PRKCB 是脂多糖免疫应答相关的候选基因,涉及 B 细胞活性和抗原应答。机制上,PRKCB 参与 B 细胞受体介导的核因子 κB 信号通路的激活[51]。杨旋等[43]利用高通量测序检测了 4 例 RA 患者及 3 例健康对照组的外周血单个核细胞,发现显著性差异改变[log2(FC)≥1.0 或 log2(FC)≤−1.0,且错误发现率≤0.05]的 circRNA 中有 41 种 circRNA 表达上调,30 种表达下调,利用 qRT-PCR 发现 hsa_circ_0000396 和 hsa_circ_0130438 在 RA 患者外周血中表达量显著下调(P<0.05),并通过 KEGG pathway 等生物信息学分析发现其可能与炎症相关的通路如肿瘤坏死因子(tumor necrosis factor,TNF)信号通路、核因子 κB 信号通路等有关,且进一步发现 hsa_circ_0000396 可能是 RA 的潜在生物标志物(AUC=0.809)。宋新强等[44]发现 hsa_circ_0005397 表达水平在 RA 患者外周血中明显低于健康对照组(P<0.016),并可能通过影响或竞争性结合 hsa_miR_125a_3p 来调节靶基因,进而参与炎症过程。
3.3 其他 circRNA
Li 等[39]研究发现,hsa_circ_0001859 可以通过直接抑制 miR-204/211 活性而上调激活转录因子 2(activating transcription factor 2,ATF2),促进 RA 的炎症过程。就机制而言,hsa_circ_0001859 通过隐藏 miR-204/211 与其靶基因 ATF2 的 3’-非翻译区(untranslated regions,UTR)结合位点,与 ATF2 竞争性结合 miR-204/211,进而上调 ATF2 的表达。该研究在 RA 滑膜细胞中进一步证实,沉默 hsa_circ_ 0001859 能使 ATF2 甚至白细胞介素(interleukin,IL)-6、TNF 和 IL-1β 表达明显下调,抑制炎症反应,ATF2 是环磷腺苷应答元件结合蛋白家族的一员,其与丝裂原活化蛋白激酶、Toll 样受体等炎症信号通路密切相关,而 IL-6、TNF 和 IL-1β 是信号通路的下游分子,与炎症活性有关。这表明 hsa_circ_0001859 在滑膜组织中可能参与了慢性炎症的过程。
CircRNA 在人外周血中存在差异表达具有区分 RA 患者与健康人的潜能[43],可见 circRNA 可能成为诊断 RA 的新型生物标志物,并为 RA 的诊断和治疗提供新的依据。
4 总结与展望
目前已有大量的 circRNA 被发现,circRNA 的功能及机制多样化,与人类疾病发生密切相关,具有作为癌症等疾病监测的新型靶标分子的潜能[52],已初步展现了其潜在的应用价值。但与 RNA 家族其他成员比,circRNA 的研究仍不够深入,circRNA 具体的生成机制还不清楚,有待进一步探索。而随着高通量测序和生物信息学的发展,人们对 circRNA 的研究将更加透彻、深入,很可能为 RA 及其他自身免疫病的发病机制、疾病发展等相关研究提供重要线索,丰富 RA 的研究内容,帮助我们建立基于 circRNA 的 RA 临床治疗和诊断策略。
环状 RNA(circular RNA,circRNA)是由前体 RNA 可变剪接产生的一类不具有游离的 5’ 端帽子和 3’ 端 poly(A)尾巴、以共价键形成环形结构的非编码 RNA 分子,且不易被核酸外切酶 RNaseR 降解[1]。1976 年人们在仙台病毒(Sendai virus)中首次发现 circRNA[2]。早期研究认为 circRNA 是一类剪接副产物或因错误剪接而形成的低丰度 RNA 分子[3],未引起关注。随着测序技术及生物信息学的发展和广泛应用,大量的 circRNA 被发现,如 Jeck 等[4]在人类成纤维细胞中检测出超过 25 000 种 circRNA;Salzman 等[5]发现了大量与人类基因表达密切相关的 circRNA。研究表明 circRNA 具有存在广泛、高度稳定、时序及疾病特异性等生物学特征,对于基因表达也起到调控作用[5-7]。
类风湿关节炎(rheumatoid arthritis,RA)是一种以慢性滑膜炎为特征的自身免疫性疾病,其发病受多因素影响,最终可能出现不同程度关节软骨和骨质破坏[8-9]。现今越来越多的报道揭示了 circRNA 与多种人类疾病有关,如血管动脉粥样硬化性疾病、癌症和自身免疫性疾病等[10-14],预示着 circRNA 可能成为新一代的生物标志物或治疗靶点。而到目前为止,还很少有研究探讨 circRNA 与 RA 之间的关系,因此本文就 circRNA 在 RA 中的最新研究进展作一综述。
1 CircRNA 的分类
目前,根据形成方式的不同,内源性 circRNA 主要分为 3 种:外显子 circRNA(exonic circRNA,ecircRNA)、内含子 circRNA(circular intronic RNA,ciRNA)以及外显子-内含子 circRNA(exonic-intronic circRNA,EI ciRNA)[15]。这些 circRNA 分子间存在竞争性平衡关系,可影响 mRNA 的表达。
1.1 ecircRNA
ecircRNA 为由外显子序列构成的 circRNA,主要存在于细胞质中[4],也是目前研究得最多的 circRNA。CircRNA 的外显子环化包括套索驱动环化和内含子配对驱动环化两种模式。前者是由前体 mRNA 在剪接过程中发生部分折叠而拉近非相邻的外显子,发生外显子跳跃的经典剪接。被跳过外显子的上、下游结合形成套索前体,随后内部剪接去除内含子,形成 ecircRNA[16]。后者是两侧内含子的反向互补序列碱基配对,诱导二者间下游剪接供体与上游剪接受体结合,而环化的外显子被释放成为了 circRNA 分子[17]。
1.2 ciRNA
内含子序列占人类基因组的 20% 以上,但人们普遍认为大多数内含子或内含子片段是不稳定的[18]。Zhang 等[19]提出了内含子来源的非编码 circRNA。该研究也提示了非编码内含子转录子在其亲本编码基因上的顺式调控作用。机体内置于细胞核中的 ciRNA 是由内含子独立环化形成,并主要源于内含子的自我剪接。ciRNA 的形成有赖于含有 3’ 剪接位点附近的 11 nt C 富集区域和邻近 5’ 端的 7 nt GU 富集元件,具有少量的微 RNA(microRNA,miRNA)靶点[20]。上述特殊结构使内含子在剪接过程中避免被脱支酶消化而形成 ciRNA[6]。
1.3 EI ciRNA
EI ciRNA 主要定位于细胞核。在 circRNA 结构中环化的外显子中间保留了内含子序列,我们称之为外显子-内含子环状结构或 EI ciRNA[21],但具体发生机制尚不明确。Li 等[18]研究发现 EI ciRNA 可通过 U1 核小 RNA 和 EI ciRNA 之间的特定的 RNA-RNA 相互作用进行转录控制的调控策略,增强亲本基因的表达。上述研究揭示了基因表达在转录及转录后水平的多样性和复杂性。
2 CircRNA 的生物学功能
2.1 MiRNA 分子海绵
CircRNA 富含与 miRNA 的结合位点,可抑制 mRNA 与靶基因相结合,从而发挥 miRNA“海绵”效应,这是目前 circRNA 被报道最多的功能。Hansen 等[22]研究发现,小脑变性相关蛋白 1 反义转录物(antisense to the cerebellar degeneration-related protein 1 transcript,CDR1as)是小脑变性相关蛋白 1(cerebellar degeneration related protein 1,CDR1)基因的环状天然反义转录物,也被称为 miRNA-7(miR-7)的 circRNA 海绵(ciRS-7),对 miR-7 起到负调控作用[23],上调其靶基因的表达水平,提示 circRNA 可通过参与到竞争的内源 RNA 网络,从而在转录水平上对靶基因进行调控来发挥生物学作用[24]。在胚胎中超高表达 CDR1as 或沉默 miR-7 可导致斑马鱼中脑缩小[25],暗示了 CDR1as 可作为 miR-7 分子海绵的作用。Bai 等[26]研究发现 circDLGAP4 可作为内源性 miRNA-143(miR-143)海绵来抑制其活性,解除 miR-143 对 HECTD1 基因表达的抑制作用,可见,circRNA 可提高靶基因的表达水平[27],当 miRNA 被 circRNA 吸附时,mRNA 翻译合成蛋白质[28]。虽然当前,circRNA 起 miRNA 的海绵作用还在肿瘤[29]、卵泡发育[30]及骨肉瘤的生长[31]等研究中也有体现,但 circRNA 是否普遍具有 miRNA 海绵作用还有待进一步验证。
2.2 CircRNA 调控基因转录
与传统线性 RNA 不同,circRNA 在真核转录组中大量存在。一些内含子来源的 circRNA 存在于细胞核中,可与 RNA 聚合酶Ⅱ转录复合体结合,以顺式作用方式对亲本基因的转录进行调控[11],进而影响靶基因表达。如 ci-ankrd52 是来源于锚蛋白重复结构域 52(ankyrin repeat domain 52,ANKRD52)基因第 2 个内含子的 circRNA,研究表明,它可参与 RNA 聚合酶Ⅱ的延伸机制,起正向调控基因的转录活性作用,而敲除 ci-ankrd52 后,ankrd52 mRNA 表达显著减少,circ-sirt7 也具有与 ci-ankrd52 相同的特征[19],表明细胞核内的 circRNA 可能参与基因的转录调控。此外,还有一些定位在细胞核中的外显子与内含子共同来源的 EI ciRNA(如 circ-EIF3J 和 circ-PAIP2),可与小核糖体 U1 snRNP 或 RNA 聚合酶Ⅱ相互作用促进亲本基因的转录[18],也暗示了 circRNA 的种类及功能具有多样性。
2.3 CircRNA 调节 RNA 结合蛋白
此前已有研究报道线性 RNA 可吸附蛋白因子[32],现研究发现 circRNA 也能与有关蛋白[主要是 RNA 结合蛋白(RNA-binding protein,RBP)]结合形成复合体,进而发挥生物学功能[17]。例如,circRNA(如 CDR1as 和 Sry)可以与 Argonaute(AGO)蛋白结合,调控 AGO mRNA 的翻译过程[33]。Ashwal-Fluss 等[16]发现作为 RBP 的剪接因子肌盲蛋白(muscle blind-like,MBL)可结合其亲本基因的外显子并促进其环化形成 circMBL,二者相互作用可降低 MBL 有效浓度,减少 circMBL 生成。
2.4 CircRNA 具有潜在翻译功能
一般情况下,很少有 circRNA 被翻译成蛋白质。但 Chen 等[34]报道了 circRNA 可启动并编码蛋白。之后 Perriman 等[35]研究出包含一个简单绿色荧光蛋白开放阅读框的 circRNA,它可以在大肠杆菌中指导绿色荧光蛋白的表达。Abe 等[36]验证了在无细胞大肠杆菌翻译系统和人类细胞中,circRNA 可通过滚动循环扩增途径有效地被翻译,并产生丰富的蛋白质产物,也表明这些在人类细胞中大量存在的 ecircRNA 所具有较大的翻译潜能。依赖 N6-甲基腺甲基化修饰[37]和依赖内部核糖体嵌入位点[38]是目前报道最多的 circRNA 两种可能的翻译机制。但 circRNA 在翻译控制中的作用仍然值得进一步研究,且目前为止尚无真核生物细胞自然产生此类具有翻译功能 circRNA 的证据。
3 CircRNA 与 RA
近些年测序结果表明 circRNA 在 RA 患者及健康人外周血单核细胞中存在表达异常,表 1 总结了与 RA 有关的 circRNA[39-45]。
3.1 高表达 circRNA
Ouyang 等[40]通过微阵列芯片技术筛选出差异表达的 circRNA 后使用实时荧光定量聚合酶链反应(real-time quantitative polymerase chain reaction,qRT-PCR)验证了 30 例 RA 患者及 25 例健康人的外周血单核细胞,发现与健康对照组相比,RA 患者中的 circRNA_092516、circRNA_003524、circRNA_103047、circRNA_104871 及 circRNA_101873 表达显著上调,其中受试者工作特征(receiver operating characteristic,ROC)曲线分析提示 circRNA_104871 对于 RA 有较高的诊断价值[ROC 曲线下面积(area under curve,AUC)=0.833,95% 置信区间(0.721,0.944),P<0.001]。进一步研究发现这些 RA 患者外周血单核细胞中具有显著差异性表达的 circRNA 与 28 处关节疾病活动度评分(Disease Activity Score 28,DAS28)、C 反应蛋白、红细胞沉降率、健康评估问卷(Health Assessment Questionnaire,HAQ)均无相关性,表明这些 circRNA 可能不是疾病活动度或者全身炎症反应的相关生物标志物。Zheng 等[41]通过微阵列芯片检测 10 例 RA 患者及 10 例健康对照者外周血单核细胞中的 cricRNA 表达情况,发现有 255 个 circRNA 表达显著上调,329 种 cricRNA 显著下调,并采用 qRT-PCR 验证了 hsa_circ_0092285、hsa_circ_ 0058794 表达显著上调。而 hsa_circ_0092285 可能通过与 PNKP 基因作用参与到 RA 的氧化应激与炎症反应中。hsa_circ_0058794 由 AGAP1 基因拼接而来,进一步研究发现它能促进滑膜细胞侵袭软骨组织[46],有可能成为 RA 患者预后评估的生物标志物。Tang 等[42]报道 ciRS-7 在 RA 患者外周血中显著高表达,并发现 RA 患者 ciRS-7 的相对表达量与血浆抗环瓜氨酸多肽抗体水平呈正相关(r=0.564 0,P=0.014 8);该研究进一步揭示哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)mRNA 在 RA 患者外周血中的表达显著高于健康对照组,并与 ciRS-7 表达呈正相关,与 miR-7 表达呈负相关,推测在 RA 患者中 ciRS-7 可能作为 miR-7 的“海绵”并“吸收”一定量的 miR-7 进而上调 mTOR 的表达。这些结果表明 ciRS-7 可能是 RA 潜在的生物标志物(AUC=0.766),为 RA 的诊断和靶向治疗提供了新的思路。
3.2 低表达 circRNA
有研究发现源自于类固醇脱氢酶样蛋白 2(hydroxysteroid dehydrogenase like 2,HSDL2)基因的 hsa_circ_0088088 表达水平在 RA 患者外周血中明显下调[41],并可能增加 RA 患者心血管事件的风险[47]。就机制而言,HSDL2 参与了脂肪酸和胆固醇的代谢和稳态的定位[48],其下调可能影响脂质分解并增加游离脂肪酸水平[49],从而增加 RA 患者动脉粥样硬化的风险[50]。与健康人相比,在 RA 患者外周血中显著下调的 hsa_circ_0038644 于 PRKCB 中编码[41],而 PRKCB 是脂多糖免疫应答相关的候选基因,涉及 B 细胞活性和抗原应答。机制上,PRKCB 参与 B 细胞受体介导的核因子 κB 信号通路的激活[51]。杨旋等[43]利用高通量测序检测了 4 例 RA 患者及 3 例健康对照组的外周血单个核细胞,发现显著性差异改变[log2(FC)≥1.0 或 log2(FC)≤−1.0,且错误发现率≤0.05]的 circRNA 中有 41 种 circRNA 表达上调,30 种表达下调,利用 qRT-PCR 发现 hsa_circ_0000396 和 hsa_circ_0130438 在 RA 患者外周血中表达量显著下调(P<0.05),并通过 KEGG pathway 等生物信息学分析发现其可能与炎症相关的通路如肿瘤坏死因子(tumor necrosis factor,TNF)信号通路、核因子 κB 信号通路等有关,且进一步发现 hsa_circ_0000396 可能是 RA 的潜在生物标志物(AUC=0.809)。宋新强等[44]发现 hsa_circ_0005397 表达水平在 RA 患者外周血中明显低于健康对照组(P<0.016),并可能通过影响或竞争性结合 hsa_miR_125a_3p 来调节靶基因,进而参与炎症过程。
3.3 其他 circRNA
Li 等[39]研究发现,hsa_circ_0001859 可以通过直接抑制 miR-204/211 活性而上调激活转录因子 2(activating transcription factor 2,ATF2),促进 RA 的炎症过程。就机制而言,hsa_circ_0001859 通过隐藏 miR-204/211 与其靶基因 ATF2 的 3’-非翻译区(untranslated regions,UTR)结合位点,与 ATF2 竞争性结合 miR-204/211,进而上调 ATF2 的表达。该研究在 RA 滑膜细胞中进一步证实,沉默 hsa_circ_ 0001859 能使 ATF2 甚至白细胞介素(interleukin,IL)-6、TNF 和 IL-1β 表达明显下调,抑制炎症反应,ATF2 是环磷腺苷应答元件结合蛋白家族的一员,其与丝裂原活化蛋白激酶、Toll 样受体等炎症信号通路密切相关,而 IL-6、TNF 和 IL-1β 是信号通路的下游分子,与炎症活性有关。这表明 hsa_circ_0001859 在滑膜组织中可能参与了慢性炎症的过程。
CircRNA 在人外周血中存在差异表达具有区分 RA 患者与健康人的潜能[43],可见 circRNA 可能成为诊断 RA 的新型生物标志物,并为 RA 的诊断和治疗提供新的依据。
4 总结与展望
目前已有大量的 circRNA 被发现,circRNA 的功能及机制多样化,与人类疾病发生密切相关,具有作为癌症等疾病监测的新型靶标分子的潜能[52],已初步展现了其潜在的应用价值。但与 RNA 家族其他成员比,circRNA 的研究仍不够深入,circRNA 具体的生成机制还不清楚,有待进一步探索。而随着高通量测序和生物信息学的发展,人们对 circRNA 的研究将更加透彻、深入,很可能为 RA 及其他自身免疫病的发病机制、疾病发展等相关研究提供重要线索,丰富 RA 的研究内容,帮助我们建立基于 circRNA 的 RA 临床治疗和诊断策略。